

魚の摂食行動は、成長と繁殖に利用するエネルギーを得るための手段です1。そのため、現在まで様々な研究がなされてきました。研究によって摂食行動は、流れ、餌密度、光の強さ、餌の大きさ、捕食者の存在など様々な要因によって変化することが分かってきました2-11。

中でも
しかし、これらの研究は主に海や川に住む泳ぐ魚を対象にしています。泳ぐことのできる魚たちは泳いで60%以上も流れを軽減すると言われる珊瑚の隙間などに隠れたりすることができます13-15。一方でチンアナゴのような珊瑚礁辺縁部に住み、巣穴以外の隠れ家を持たない魚は流れに対処した独特な摂食様式をもつことが想像されます。
このようなことから私たちはチンアナゴを対象として、流速が与える摂食行動への影響を明らかにすることを目的として研究をしています。チンアナゴ(Gorgasia sillneriという種)の流速と摂食行動の関係はKhrizmanら(2018)に紅海で初めて研究されました16。研究では高流速下でも摂食率が落ちず、シミュレーションによって流速に対して姿勢を変化させることで抵抗を大幅に減らしていることが分かりました。
ただし、この研究はフィールドで実施されたものであり、流速以外の影響を排除できないなどの理由から、実験下での研究が待たれました。
私たちは流速を自由に変化させることができる「フルーム」と呼ばれる下の画像のような水槽を使うことでチンアナゴ(英名:spotted garden eel, 学名:Heteroconger hassi)の摂食行動に与える流速の影響を実験下で詳細に調べています。
研究は様々な分野の研究者が集まる沖縄科学技術大学院大学(OIST)で行われ、「流れ」という物理的要素が与える「チンアナゴの行動」という生物への影響を調べる

時間あたりにどれくらいの数のプランクトンを食べたかを表す
行動の3次元再構築は体のトラッキングと3D再構成の2つから成ります。行動学の世界ではトラッキングはビーズなど目立つもので動物を標識して、それらをソフトウェアを使って追跡するのが主流でしたが、チンアナゴを標識するのは容易でない上、標識そのものの影響も考慮しなければいけません。私たちの研究では撮影したビデオの中でチンアナゴの特徴的な点(目や大きい黒点など)を1フレーム1フレーム追跡します。これは手動で行うには途方もない作業になります。そこで
人間の目は2つあることで視覚情報を3次元に捉えています。3D再構成では複数のカメラをキャリブレーションすることで、それぞれのカメラの位置情報などを捉え、こうしてそれぞれのカメラで追跡した点を3次元空間に再構成します。こちらはMatlabのdltdvを使用しています19。
ちなみに私たちの研究ではなんとiPhoneのカメラを使用して動画撮影をしました!

このように行動の3次元再構築を行うと、餌を追跡した時間、距離、速度、角度、軌跡など様々なパラメーターを詳細に調べることができます。これらのパラメーターは採餌モデルを作成する際にも重要になります。こうした詳細な行動解析によって、
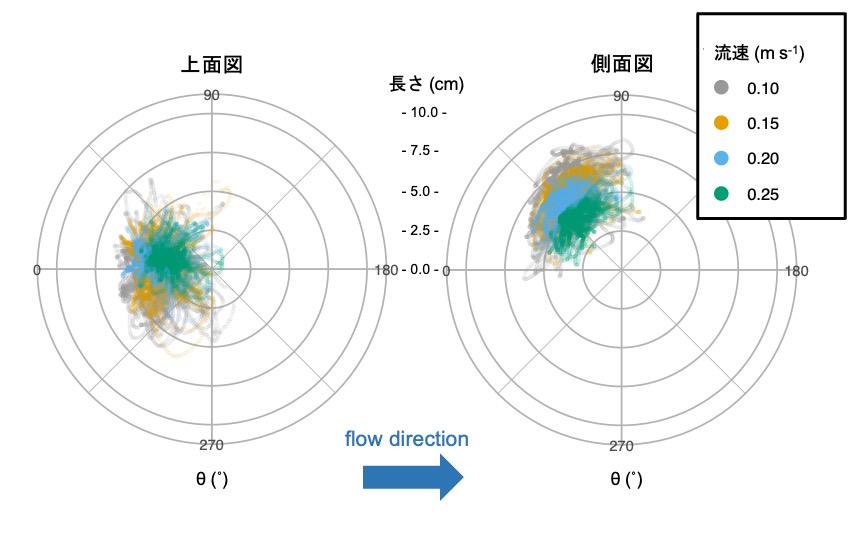
このグラフは流速を変化させた時のチンアナゴの顔の動きを極座標で表したものです。点は1フレームごとの顔の位置を表しています。これを見ると、
チンアナゴの流速への応答についてさらに詳しく知りたい方は、こちらの
Meet the garden eel – a shy creature who almost never leaves its burrow, even to feed. Now, the OIST Marine Biophysics Unit has conducted the first lab study on garden eels, revealing how their lifestyle pays off when feeding in strong currents @J_Exp_Biol https://t.co/RMxIyZoy9r pic.twitter.com/dxENPFuiEU
— OIST (@OISTedu) July 21, 2022
これまで話したように、チンアナゴ含め水中で生きる魚たちにとって、水の流れは彼らの行動や代謝に影響を与える大切な環境の一つです。特に流れの速さが魚類に与える影響はよく研究されており、魚たちは流速によって泳ぎ方や餌を食べる行動を変化させたり、消費エネルギーが変化したり、岩陰に隠れるような行動を取ったりします。5,12,13,20,21

これまでの多くの研究は、平均の流れの速さ(平均流速)の影響を調べていましたが、近年になって
しかし、これらの研究では流れのパターンが明確であるケースが多く、魚たちが流れのパターンを学習したり予測することが可能だと考えられます。そこで私たちの研究ではこのような特定のパターンが明らかでなく、不規則な流れである

乱流の影響を実験室で調べるためには、魚たちが実際の環境でどのくらいの強さの乱流を経験しているか調べる必要があります。そこではじめに、

これらの流れの条件下で、餌を食べる行動をデバスズメダイとチンアナゴを対象に調べました。すると、
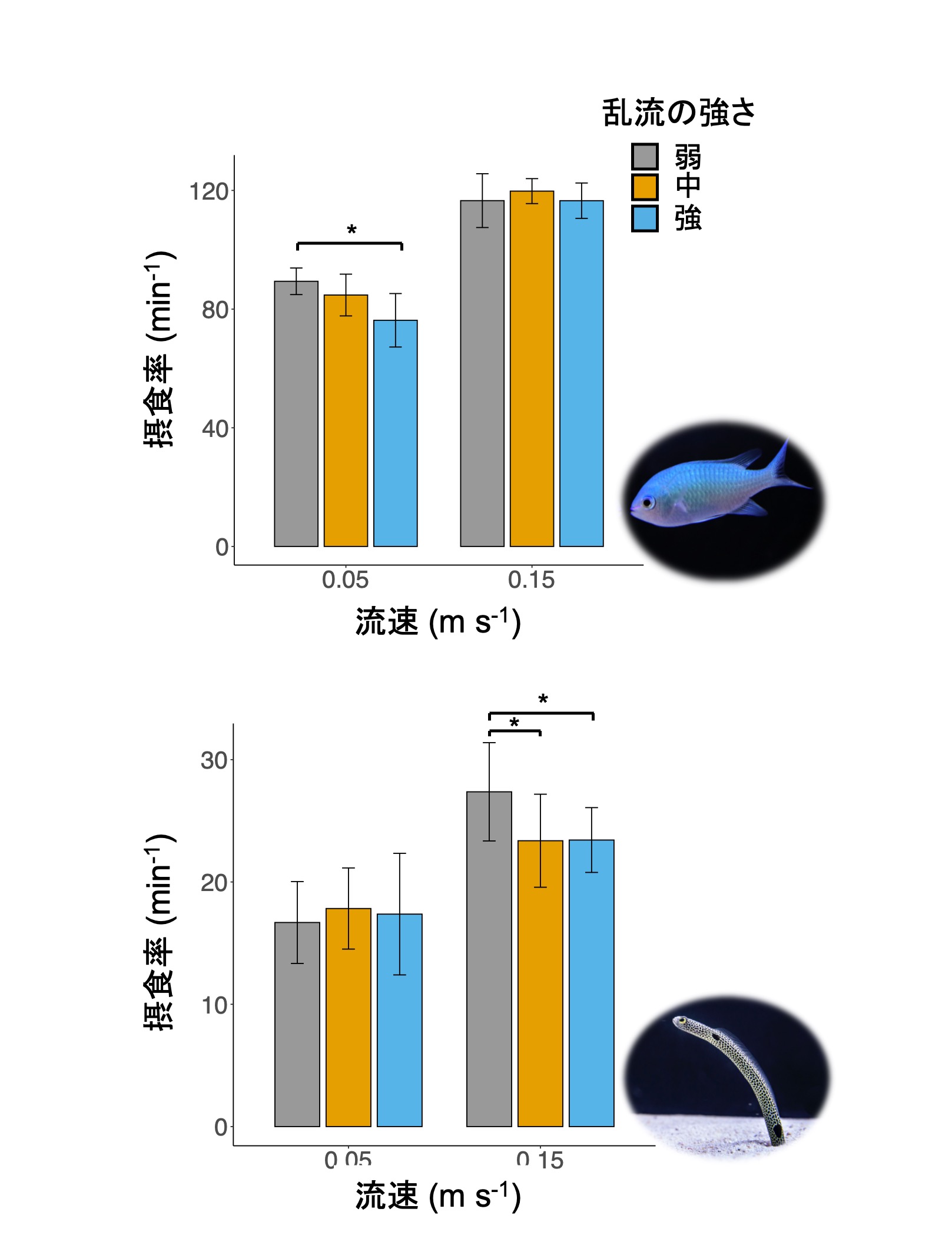
これらの結果は、それぞれの種の生息地の流れに対する適応を示していると考えられました。
チンアナゴとデバスズメダイの乱流への応答についてさらに詳しく知りたい方は、私たちが書いた



~参考文献~
1. Stoner, A. W. Effects of environmental variables on fish feeding ecology: Implications for the performance of baited fishing gear and stock assessment. J. Fish Biol. 65, 1445–1471 (2004).
2. Clarke, R. D., Finelli, C. M. & Buskey, E. J. Water flow controls distribution and feeding behavior of two co-occurring coral reef fishes: II. Laboratory experiments. Coral Reefs 28, 475–488 (2009).
3. Finelli, C. M., Clarke, R. D., Robinson, H. E. & Buskey, E. J. Water flow controls distribution and feeding behavior of two co-occurring coral reef fishes: I. Field measurements. Coral Reefs 28, 461–473 (2009).
4. Fulton, C. J., Bellwood, D. R. & Wainwright, P. C. Wave energy and swimming performance shape coral reef fish assemblages. Proc. R. Soc. B Biol. Sci. 272, 827–832 (2005).
5. Kiflawi, M. & Genin, A. Prey flux manipulation and the feeding rates of reef-dwelling planktivorous fish. Ecology 78, 1062–1077 (1997).
6. Noda, M., Kawabata, K., Gushima, K. & Kakuda, S. Importance of zooplankton patches in foraging ecology of the planktivorous reef fish Chromis chrysurus (Pomacentridae) at Kuchinoerabu Island, Japan. Mar. Ecol. Prog. Ser. 87, 251–263 (1992).
7. Hill, J. & Grossman, G. D. An Energetic Model of Microhabitat Use for Rainbow Trout and Rosyside Dace. Ecology 74, 685–698 (1993).
8. Manatunge, J. & Asaeda, T. Optimal foraging as the criteria of prey selection by two centrarchid fishes. Hydrobiologia 391, 223–240 (1998).
9. Howard, E. W. & Bori, L. O. Behavior of Marine Animals: Current Perspectives in Research. Plenum Press New York-London vol. 2 (Plenum Press, New York, 1972).
10. Rickel, S. & Genin, A. Twilight transitions in coral reef fish: The input of light-induced changes in foraging behaviour. Anim. Behav. 70, 133–144 (2005).
11. Morgan, M. J. The influence of hunger, shoal size and predator presence on foraging in bluntnose minnows. Anim. Behav. 36, 1317–1322 (1988).
12. Heatwole, S. J. & Fulton, C. J. Behavioural flexibility in reef fishes responding to a rapidly changing wave environment. Mar. Biol. 160, 677–689 (2013).
13. Johansen, J., Bellwood, D. & Fulton, C. Coral reef fishes exploit flow refuges in high-flow habitats. Mar. Ecol. Prog. Ser. 360, 219–226 (2008).
14. Johansen, J. L., Fulton, C. J. & Bellwood, D. R. Avoiding the flow: Refuges expand the swimming potential of coral reef fishes. Coral Reefs 26, 577–583 (2007).
15. Taguchi, M. & Liao, J. C. Rainbow trout consume less oxygen in turbulence: the energetics of swimming behaviors at different speeds. J. Exp. Biol. 214, 1428–1436 (2011).
16. Khrizman, A., Ribak, G., Churilov, D., Kolesnikov, I. & Genin, A. Life in the flow: unique adaptations for feeding on drifting zooplankton in garden eels. J. Exp. Biol. 221, (2018).
17. Mathis, A. et al. DeepLabCut: markerless pose estimation of user-defined body parts with deep learning. Nat. Neurosci. 21, 1281–1289 (2018).
18. Nath, T. et al. Using DeepLabCut for 3D markerless pose estimation across species and behaviors. Nat. Protoc. 14, 2152–2176 (2019).
19. Hedrick, T. L. Software techniques for two- and three-dimensional kinematic measurements of biological and biomimetic systems. Bioinspiration and Biomimetics 3, (2008).
20. O’Brien, W. J., Barfield, M. & Sigler, K. The functional response of drift-feeding Arctic grayling: the effects of prey density, water velocity, and location efficiency. Can. J. Fish. Aquat. Sci. 58, 1957–1963 (2001).
21. Piccolo, J. J., Hughes, N. F. & Bryant, M. D. Water velocity influences prey detection and capture by drift-feeding juvenile coho salmon ( Oncorhynchus kisutch) and steelhead ( Oncorhynchus mykiss irideus ). Can. J. Fish. Aquat. Sci. 65, 266–275 (2008).
22. Liao, J. C., Beal, D. N., Lauder, G. V. & Triantafyllou, M. S. Fish Exploiting Vortices Decrease Muscle Activity. Science 302, 1566–1569 (2003).
23. Liao, J. C. A review of fish swimming mechanics and behaviour in altered flows. Phil. Trans. R. Soc. B 362, 1973–1993 (2007).